
Page 47 - 《华中农业大学学报》2020年第6期
P. 47
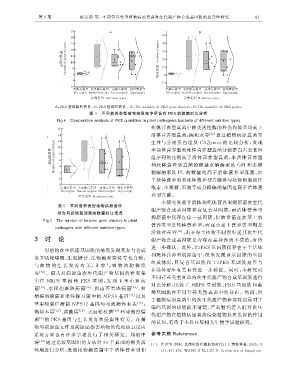
第 6 期 祝友朋 等:不同营养类型植物病原真菌次生代谢产物合成基因簇的差异性研究 4 1
A : PKS基因簇的数量; B : PKS基因的数量. A : ThenumberofPKSg eneclusters ; B : ThenumberofPKSg enes.
图 4 不同营养类型植物病原菌中所含有 PKS 的数量对比分析
Fi g .4 Com p arativeanal y sisofPKSq uantitiesinp lant p atho g enicbacteriaofdifferentnutritiont yp es
死体营养型真菌中糖类活性酶的种类和数量均高于
活体营养型真菌; 陈相永等 [ 25 ] 通过植物病原真菌寄
生性与分泌蛋白组及 CAZ y mes 的比较分析, 发现
半活体营养型和死体营养型真菌分泌蛋白占全基因
组序列的比例高于活体营养型真菌, 半活体营养型
和死体营养型真菌的糖基水解酶家族 GH 和多糖
裂解酶家族 PL 的数量均高于活体营养型真菌, 且
半活体营养型和死体营养型真菌参与植物细胞壁纤
维素、 木聚糖、 果胶等成分降解的基因也高于活体营
养型真菌.
本研究发现半活体和死体营养型病原菌次生代
图 5 不同营养类型植物病原菌中
谢产物合成基因簇存在复合基因簇, 而活体营养型
所含有萜烯基因簇的数量对比情况
病原菌中仅存在单一基因簇, 但核盘菌在油菜上的
Fi g .5 Thenumberofter p eneg eneclustersinp lant
p atho g enswithdifferentnutritiont yp es 营养类型是死体营养型, 而在小麦上营养类型则是
活体营养型 [ 26 ] , 由于寄主植物不同而引起其次生代
3 讨 论 谢产物合成基因簇是否存在差异性尚不清楚, 有待
进一步确认.此外, T3PKS 基因簇仅存在于半活体
对植物次生代谢基因簇的研究发现其参与合成
和死体营养型病原菌中, 但未发现该基因簇的基因
苯并噁唑嗪酮、 生氰糖苷、 生物碱和萜烯等化合物,
组成情况, 其复合基因簇和 T3PKS 基 因簇是否与
与 植 物 的 生 长 发 育 有 关, 并 参 与 植 物 的 防 御 反
非活体寄生有关有待进一步验证.同时, 本研究对
应 [ 20 ] .前人对病原菌次生代谢产物基因的研究集
不同营养类型真菌的次生代谢产物合成基因簇进行
中在 NRPS 基 因 和 PKS 基 因, 发 现 玉 米 小 斑 病
对比分析, 比较了 NRPS 基因簇、 PKS 基因簇和萜
菌 [ 15 ] 、 水稻胡麻斑病菌 [ 16 ] 、 拟南芥黑斑病菌 [ 16 ] 、 柑
烯基因簇在不同营养类型真菌中的分布.然而, 对
橘褐斑病菌和禾谷镰刀菌中的 NPS6 基因 [ 17 ] 以及
于植物病原真菌中的次生代谢产物研究特别是对于
苹果树腐烂病菌 NPS12 基因均与致病性有关 [ 18 ] ;
编码基因的功能尚不清楚, 严重制约着人们对次生
渐绿木霉 [ 21 ] 、 黄曲霉 [ 22 ] 、 大丽轮枝菌 [ 23 ] 和球孢白僵
代谢产物在植物病原真菌侵染植物后所发挥的作用
菌 [ 5 ] 的 PKS 基因与生长发育和侵染性有关.在植
的认知, 有待于今后开展相关生物学试验研究.
物与病原菌互作及病原菌损害植物的免疫防卫反应
系统方面也有许多学者进行了相关研究.如赵中 参考文献 References
涛 [ 24 ] 通过比较基因组的方法对 94 个真菌的糖类活
[ 1 ] 汪世华, 刘阳 . 真菌次级代谢的新时代[ J ] . 菌物学报, 2020 , 39
性酶进行分析, 发现植物病原菌中半活体营养型和 ( 3 ): 471G476.WANGS H , LIU Y.Aneweraoffun g alsecG